

Chemical properties of leaf litter among tree species with different mycorrhizal types in southern subtropical China
-
摘要: 文章以南亚热带24种木本植物(丛枝菌根、外生菌根和固氮树种各8种)叶凋落物为研究对象,探讨不同菌根类型树种叶凋落物化学性质的差异性。结果表明:叶凋落物在碳相关化学性质、养分含量以及化学计量比特征方面均表现出较大程度变异。不同菌根类型树种碳相关化学性质没有显著差异(P > 0.05),而养分含量(N、P、Mg)和化学计量特征(C/P、N/P)差异显著(P < 0.01)。固氮树种叶凋落物N含量(16.1 g · kg−1)显著高于丛枝菌根树种(10.1 g · kg−1)和外生菌根树种(10.7 g · kg−1),但丛枝菌根树种叶凋落物P含量(0.4 g · kg−1)要显著高于固氮树种(0.1 g · kg−1)和外生菌根树种(0.08 g · kg−1),而外生菌根树种Mg含量(0.51 g · kg−1)则显著高于丛枝菌根树种(0.19 g · kg−1)和固氮树种(0.03 g · kg−1);此外,固氮树种C/P和N/P均显著高于丛枝菌根树种。在南亚热带退化生态系统恢复过程中,应考虑不同菌根类型树种叶凋落物化学性质的差异,依据不同菌根类型树种养分利用策略的差异性选择适宜的恢复树种。Abstract:
The sample plot is located in the Experimental Center of Tropical Forestry, Chinese Academy of Forestry, the southwest of Guangxi, China. In a typical southern subtropical monsoon climate, the annual temperature of this area averages at 21 ℃ with the lowest average at 12.5 ℃ in January and the highest at 27 ℃ in July. The average annual precipitation is between 1,200 to 1,500 mm, and shows obvious seasonality with the wet season from April to September and the dry season from October to March in the following year. The area is characterized by a typical landform with peak-cluster depression at the elevation ranging from 150 to 800 m. The main soil type in this area is red soil developed from carbonatite. Taking the leaf litter of 24 tree species with different mycorrhizal types (respective eight species for arbuscular mycorrhizal fungi, ectomycorrhizal fungi and nitrogen-fixers) as the research object, we measured the chemical properties of leaf litter (non-structure carbohydrate, water-soluble carbon, lignin, cellulose, hemicellulose and tannin) and nutrient concentration (N, P, K, Ca, Mg and Mn), analyzed the relationships of these properties, and explored the effects of different mycorrhizal types on the chemical properties. The results indicate that the chemical properties of leaf litter exhibited a fair degree of variation in carbon-related properties, nutrient contents and stoichiometric properties. There were no significant differences in carbon-related chemical properties among tree species with different mycorrhizal types (P>0.05). However, the nutrient content (N, P and Mg) and stoichiometry properties of leaf litter (C/P and N/P) were significantly different among mycorrhizal types. Nitrogen-fixer trees showed a significantly higher leaf litter N (16.1 g·kg−1) than the trees with arbuscular mycorrhizal (10.1 g·kg−1) and ectomycorrhizal fungi (10.7 g·kg−1). However, tree species with arbuscular mycorrhizal fungi (0.4 g·kg−1) exhibited a significantly higher P concentration than the trees with ectomycorrhizal fungi (0.1 g·kg−1) and N-fixer (0.08 g·kg−1) trees. The concentration of Mg (0.51 g·kg−1) in the tree species with ectomycorrhizal fungi was significantly higher than the trees with arbuscular mycorrhizal fungi (0.19 g·kg−1) and N-fixer trees (0.03 g·kg−1). In addition, the C/P and N/P of N-fixer trees were significantly higher than the tree species with arbuscular. Hence the differences in the chemical properties of leaf litter among different mycorrhizal types should be considered during the restoration of degraded ecosystems, and appropriate tree species for restoration will be selected according to the strategies of nutrient utilization. -
Key words:
- litter decomposition /
- mycorrhiza /
- stoichiometry /
- carbon cycle /
- ecological restoration
-

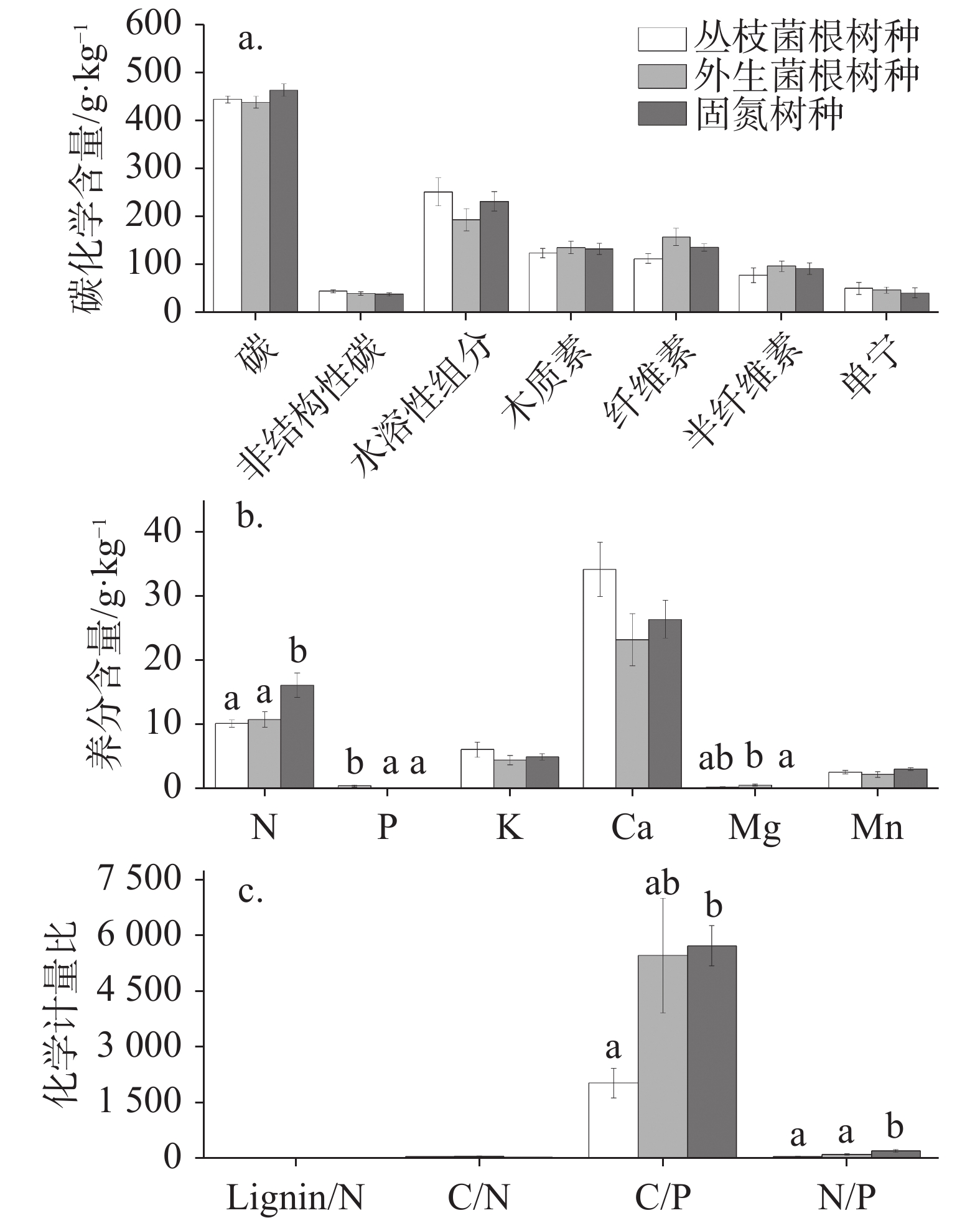
图 1 不同菌根类型树种叶凋落物化学性质
注:不同小写字母表示不同菌根树种间叶凋落物化学性质差异显著(P<0.05)。
Figure 1. Chemical properties of leaf litter among different mycorrhizal types
Note: Different small letters represent significant difference in chemical properties of leaf litter among different mycorrhizal types(P<0.05).
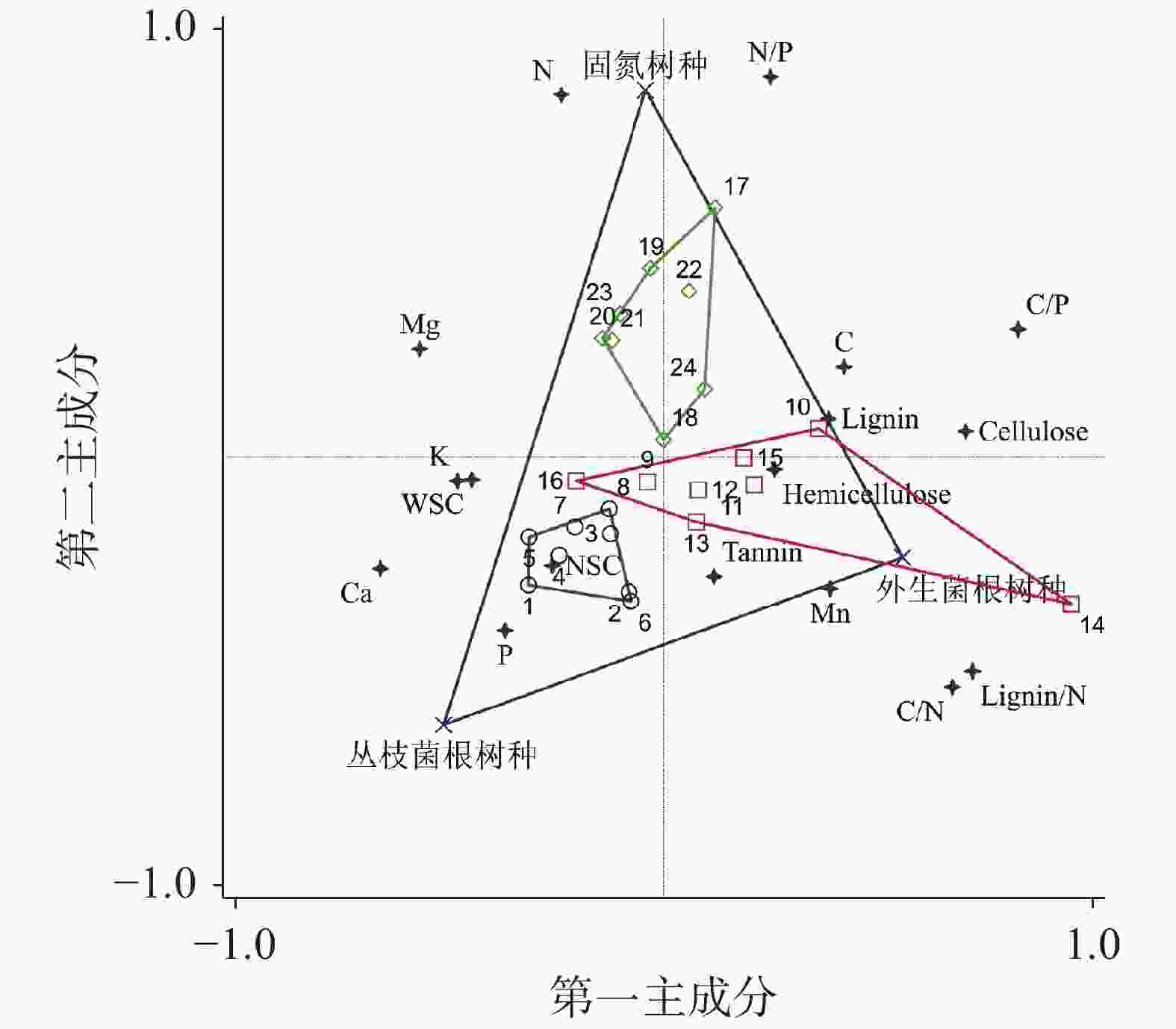
表 1 不同菌根类型树种
Table 1. Tree species with different mycorrhizal types
菌根类型 编号 物种 所属科 生活型 丛枝菌根 1 割舌树 Walsura robusta 楝科 Meliaceae 常绿阔叶 2 南酸枣 Choerospondias axillaris 漆树科 Anacardiaceae 落叶阔叶 3 东京桐 Deutzianthus tonkinensis 大戟科 Euphorbiaceae 落叶阔叶 4 望天树 Parashorea chinensis 龙脑香科 Dipterocarpaceae 常绿阔叶 5 光皮梾木 Cornus wilsoniana wanger 山茱萸科Cornaceae 落叶阔叶 6 小叶红光树 Knema globularia 肉豆蔻科 Myristicaceae 落叶阔叶 7 麻楝 Chukrasia tabularis 楝科 Meliaceae 落叶阔叶 8 山牡荆 Vitex quinata 唇形科 Lamiaceae 落叶阔叶 外生菌根 9 蚬木 Excentrodendron tonkinense 锦葵科 Malvaceae 常绿阔叶 10 青冈栎 Cyclobalanopsis glauca 壳斗科 Fagaceae 常绿阔叶 11 栓皮栎 Quercus variabilis 壳斗科 Fagaceae 常绿阔叶 12 美国山核桃 Carya illinoinensis 胡桃科 Juglandaceae 落叶阔叶 13 枫香树 Liquidambar formosana 金缕梅科Hamamelidaceae 落叶阔叶 14 湿地松 Pinus elliottii 松科 Pinaceae 常绿针叶 15 红锥 Castanopsis hystrix 壳斗科 Fagaceae 常绿阔叶 16 海南椴 Hainania trichosperma 椴树科Tiliaceae 落叶阔叶 固氮树种 17 格木 Erythrophleum fordii 豆科 Fabaceae 常绿阔叶 18 仪花 Lysidice rhodostegia 豆科 Fabaceae 常绿阔叶 19 任豆 Zenia insignis 豆科 Fabaceae 落叶阔叶 20 海红豆 Adenanthera pavonina 豆科 Fabaceae 落叶阔叶 21 顶果树 Acrocarpus fraxinifolius 豆科 Fabaceae 落叶阔叶 22 台湾相思 Acacia confusa 豆科 Fabaceae 常绿阔叶 23 降香黄檀 Dalbergia odorifera 豆科 Fabaceae 常绿阔叶 24 银合欢 Leucaena leucocephala 豆科 Fabaceae 常绿阔叶  下载: 导出CSV
下载: 导出CSV
表 2 24种叶凋落物化学性质
Table 2. Chemical properties of 24 types of leaf litter
叶凋落物化学性质/g·kg−1 均值±标准误差 最小值 最大值 变异系数(CV)/% C化学性质 碳 (C) 448.19 ± 6.46 390.66 524.45 6.92 非结构性碳 (NSC) 40.57 ± 1.94 19.28 56.27 22.91 水溶性组分 (WSC) 224.96 ± 14.40 134.09 375.59 30.70 木质素 (Lignin) 130.14 ± 6.27 93.58 182.76 23.12 纤维素 (Cellulose) 134.63 ± 7.99 70.92 229.65 28.47 半纤维素 (Hemicellulose) 88.02 ± 7.25 29.61 170.57 39.51 单宁 (Tannin) 45.48 ± 5.53 9.45 117.72 58.30 养分含量 氮 (N) 12.30 ± 0.92 3.45 22.83 35.79 磷 (P) 0.19 ± 0.06 0.03 1.38 138.75 钾 (K) 5.09 ± 0.48 0.95 11.14 45.64 钙 (Ca) 27.93 ± 2.37 6.23 50.67 39.66 镁 (Mg) 0.24 ± 0.07 0.01 1.06 130.77 锰 (Mn) 2.51 ± 0.19 0.71 3.69 36.33 化学计量特征 木质素/氮 (Lignin/N) 12.45 ± 1.70 6.23 47.84 65.50 碳/氮 (C/N) 42.73 ± 4.94 19.87 144.54 55.44 碳/磷 (C/P) 4 399.19 ±639.16 343.64 15350.37 69.68 氮/磷 (N/P) 117.87 ± 17.88 8.74 382.19 72.70
下载: 导出CSV
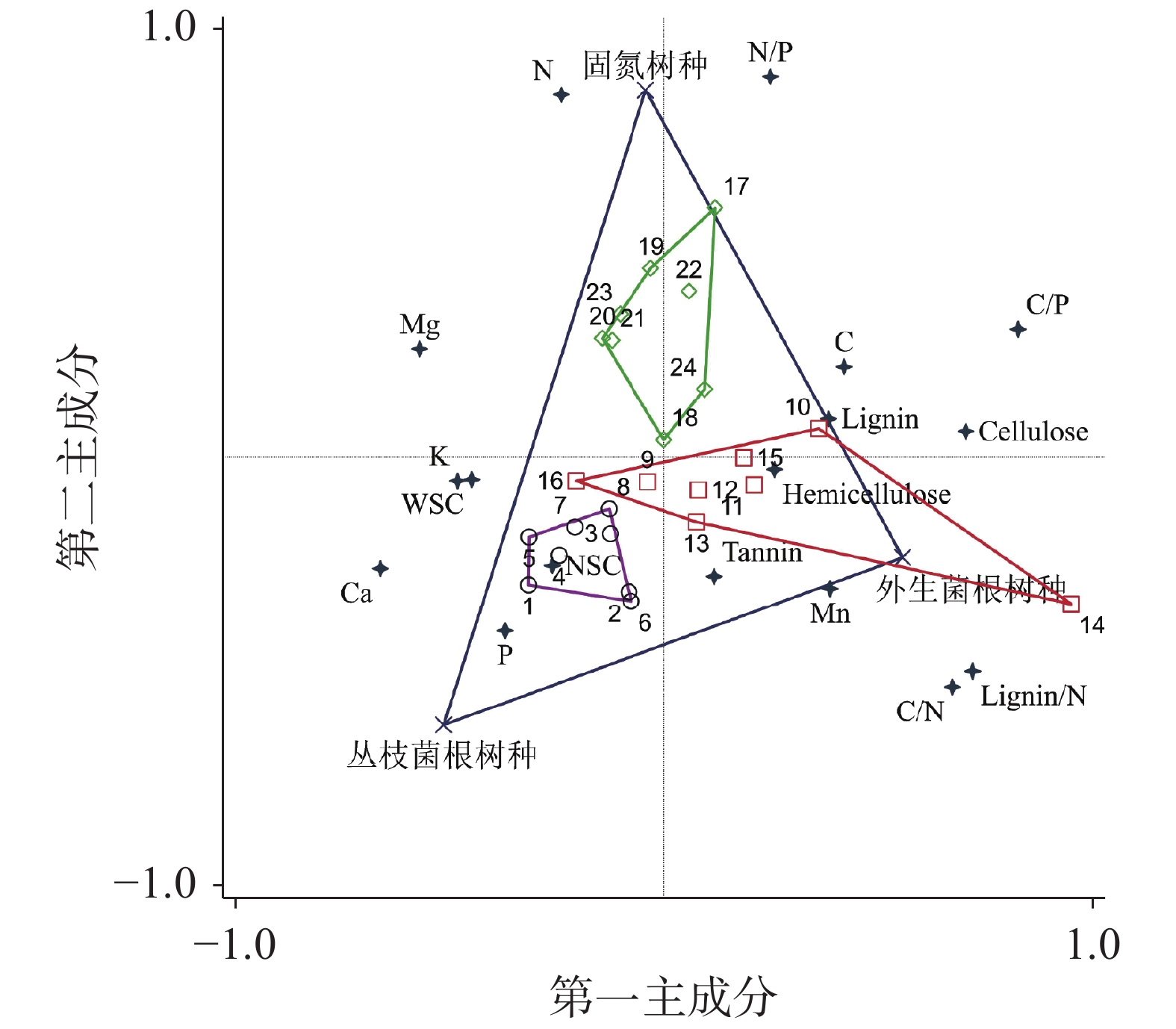
表 3 叶凋落物17种化学性质之间的相关性
Table 3. Pearson correlations of 17 chemical properties of leaf litter
化学性质 C NSC WSC Lignin Cellulose Hemi-
celluloseTannin N P K Ca Mg Mn Lignin/N C/N C/P N/P C 1 NSC −0.07 1 WSC −0.34 0.42* 1 Lignin 0.30 0.13 −0.15 1 Cellulose 0.29 −0.49* −0.55** −0.13 1 Hemicellulose −0.04 0.31 0.11 0.17 0.04 1 Tannin 0.06 −0.08 −0.05 0.12 −0.08 0.02 1 N 0.04 0.00 0.02 0.18 −0.11 −0.07 −0.24 1 P −0.22 0.28 0.11 −0.16 −0.33 −0.18 0.03 −0.12 1 K 0.02 0.04 −0.07 −0.27 −0.24 0.06 −0.14 0.05 0.38 1 Ca −0.41* 0.10 0.20 −0.32 −0.42* −0.35 0.10 −0.18 0.34 −0.20 1 Mg −0.09 −0.05 −0.37 0.25 0.30 −0.10 0.06 −0.22 −0.01 −0.30 −0.08 1 Mn −0.14 0.27 0.12 −0.23 −0.44* 0.02 0.09 0.22 0.11 0.15 0.58 * −0.34 1 Lignin/N 0.25 0.08 −0.14 0.46* 0.13 0.23 0.32 −0.75** −0.12 0.28 −0.37 −0.30 −0.13 1 C/N 0.36 0.07 −0.14 0.13 0.40 0.27 0.25 −0.74** −0.24 0.10 −0.36 −0.30 −0.21 0.87** 1 C/P 0.48* −0.13 −0.24 0.23 0.56** 0.22 −0.03 −0.04 −0.83** 0.05 −0.28 −0.41* −0.52** 0.37 0.59** 1 N/P 0.25 −0.18 −0.13 0.20 0.17 −0.06 −0.17 0.74** −0.66** −0.09 0.06 −0.22 −0.37 −0.42 −0.31 0.56** 1 注:表中数据为Pearson相关系数;*P<0.05,**P<0.01。
Note: The data in the table are Pearson correlation coefficient;* P <0.05, ** P <0.01.
下载: 导出CSV
-
[1] Lim P O, Kim H J, Nam G H. Leaf Senescence[J]. Annual Review of Plant Biology, 2007, 58:115-36. doi: 10.1146/annurev.arplant.57.032905.105316 [2] Griffiths H M, Ashton L A, Parr C L, Eggleton P. The impact of invertebrate decomposers on plants and soil[J]. New Phytologist, 2021, 231(6):2142-2149. doi: 10.1111/nph.17553 [3] 张婷, 代群威, 邓远明, 李琼芳, 董发勤, LI Bowen, Bruce W Fouke, 李相邑. 九寨沟优势植物凋落物叶片淋溶的碳氮磷释放特征[J]. 中国岩溶, 2021, 40(1):133-139.ZHANG Ting, DAI Qunwei, DENG Yuanming, LI Qiongfang, DONG Faqin, LI Bowen, Bruce W Fouke, LI Xiangyi. Release characteristics of carbon, nitrogen and phosphorus from withered leaves of dominant plants in Jiuzhaigou valley[J]. Carsologica Sinica, 2021, 40(1):133-139. [4] Hobbie S E. Plant species effects on nutrient cycling: Revisiting litter feedbacks[J]. Trends in Ecology & Evolution, 2015, 30(6):357-363. [5] Guo L L, Deng M F, Yang S, Liu W X, Wang X, Wang J, Liu L L. The coordination between leaf and fine root litter decomposition and the difference in their controlling factors[J]. Global Ecology & Biogeography, 2021, 30(11):2286-2296. [6] Bradford M A, Veen G F, Bonis A, Bradford E M, Classen A T, Cornelissen J H C, Crowther T W, De Long J R, Freschet G T, Kardol P, Manrubia-Freixa M, Maynard D S, Newman G S, Logtestijn R S P, Viketoft M, Wardle D A, Wieder W R, Wood S A, van der Putten W H. A test of the hierarchical model of litter decomposition[J]. Nature Ecology & Evolution, 2017, 1(12):1836-1845. [7] Prieto I, Almagro M, Bastida F, Querejeta J I. Altered leaf litter quality exacerbates the negative impact of climate change on decomposition[J]. Journal of Ecology, 2019, 107(5):2364-2382. doi: 10.1111/1365-2745.13168 [8] 涂春艳, 黄炳惠, 杨慧, 白冰, 莫碧琴, 曹建华. 桂林毛村岩溶区与碎屑岩区檵木和马尾松凋落叶分解研究[J]. 中国岩溶, 2018, 37(3):379-387.TU Chunyan, HUANG Binghui, YANG Hui, BAI Bing, MO Biqin, CAO Jianhua. Study on leaf litter decomposition of Loropetalum chinense andPinus massoniana in karst areas and clastic rock areas in Maocun, Guilin[J]. Carsologica Sinica, 2018, 37(3):379-387. [9] See C R, McCormack M L, Hobbie S E, Flores-Moreno H, Silver WL, Kennedy PG. Global patterns in fine root decomposition: Climate, chemistry, mycorrhizal association and woodiness[J]. Ecology Letters, 2019, 22(6):946-953. doi: 10.1111/ele.13248 [10] 金文豪, 邵帅, 陈俊辉, 秦华. 不同类型菌根对土壤碳循环的影响差异研究进展[J]. 浙江农林大学学报, 2021, 38(5):953-962. doi: 10.11833/j.issn.2095-0756.20210531JIN Wenhao, SHAO Shuai, CHEN Junhui, QIN Hua. Research progress in the impact of different mycorrhizal types on soil carbon cycling[J]. Journal of Zhejiang A&F University, 2021, 38(5):953-962. doi: 10.11833/j.issn.2095-0756.20210531 [11] Steidinger B S, Crowther T W, Liang J, van Nuland M E, Werner G D A, Reich P B, Nabuurs G J, de-Miguel S, Zhou M, Picard N, Herault B, Zhao X, Zhang C, Routh D, Consortium G F B I, Peay K G. Climatic controls of decomposition drive the global biogeography of forest-tree symbioses[J]. Nature, 2019, 569(7756):404-408. doi: 10.1038/s41586-019-1128-0 [12] Tedersoo L, Bahram M. Mycorrhizal types differ in ecophysiology and alter plant nutrition and soil processes[J]. Biological Reviews, 2019, 94(5):1857-1880. [13] Craig M E, Turner B L, Liang C, Clay K, Johnson D J, Phillips R P. Tree mycorrhizal type predicts within-site variability in the storage and distribution of soil carbon and nitrogen[J]. Global Change Biology, 2018, 24(8):3317-3330. doi: 10.1111/gcb.14132 [14] Dovrat G, Sheffer E. Symbiotic dinitrogen fixation is seasonal and strongly regulated in water-limited environments[J]. New Phytologist, 2019, 221(4):1866-1877. doi: 10.1111/nph.15526 [15] Keller A B, Phillips R P. Leaf litter decay rates differ between mycorrhizal groups in temperate, but not tropical forests[J]. New Phytologist, 2019, 222(1):554-562. [16] Averill C, Turner B J, Finzi A C. Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage[J]. Nature, 2014, 505(7484):543-545. doi: 10.1038/nature12901 [17] 何兴元, 赵淑清, 杨思河, 田春杰. 固氮树种在混交林中的作用研究Ⅲ. 固氮树种凋落物分解及N的释放[J]. 应用生态学报, 1999, 10(4):404-406. doi: 10.3321/j.issn:1001-9332.1999.04.006HE Xingyuan, ZHAO Shuqing, YANG Sihe, TIAN Chunjie. Role of nitrogen-fixing trees in mixed forest Ⅲ. leaf litter decomposition and its N release of nitrogen-fixing tree species[J]. Chinese Journal of Applied Ecology, 1999, 10(4):404-406. doi: 10.3321/j.issn:1001-9332.1999.04.006 [18] 王克林, 岳跃民, 陈洪松, 吴协保, 肖峻, 祁向坤, 张伟, 杜虎. 喀斯特石漠化综合治理及其区域恢复效应[J]. 生态学报, 2019, 39(20):7432-7440.WANG Kelin, YUE Yuemin, CHEN Hongsong, WU Xiebao, XIAO Jun, QI Xiangkun, ZHANG Wei, DU Hu. The comprehensive treatment of karst rocky desertification and its regional restoration effects[J]. Acta Ecologica Sinica, 2019, 39(20):7432-7440. [19] 王聪, 伍星, 傅伯杰, 韩兴国, 陈亚宁, 王克林, 周华坤, 冯晓明, 李宗善. 重点脆弱生态区生态恢复模式现状与发展方向[J]. 生态学报, 2019, 39( 20) : 7333-7343.WANG Cong, WU Xing, FU Bojie, HAN Xingguo, CHEN Yaning, WANG Kelin, ZHOU Huakun, FENG Xiaoming, LI Zongshan. Ecological restoration in the key ecologically vulnerable regions: Current situation and development direction[J]. Acta Ecologica Sinica, 2019, 39(20): 7333-7343. [20] Wang K L, Zhang C H, Chen H S, Yue Y M, Zhang W, Zhang M Y, Qi X K, Fu Z Y. Karst landscapes of China: Patterns, ecosystem processes and services[J]. Landscape Ecology, 2019, 34(3):2743-2763. [21] 蒋忠诚, 罗为群, 童立强, 程洋, 杨奇勇, 吴泽燕, 梁建宏. 21世纪西南岩溶石漠化演变特点及影响因素[J]. 中国岩溶, 2016, 35(5):461-468.JIANG Zhongcheng, LUO Weiqun, TONG Liqiang, CHENG Yang, YANG Qiyong, WU Zeyan, LIANG Jianhong. Evolution features of rocky desertification and influence factors in karst areas of Southwest China in the 21st century[J]. Carsologica Sinica, 2016, 35(5):461-468. [22] 欧阳帅, 项文化, 陈亮, 曾叶霖, 胡彦婷, 雷丕锋, 方晰, 邓湘雯. 南方山地丘陵区森林植被恢复对水土流失调控机制[J]. 水土保持学报, 2021, 35(5):1-9. doi: 10.13870/j.cnki.stbcxb.2021.05.001OUYANG Shuai, XIANG Wenhua, CHEN Liang, ZENG Yelin, HU Yanting, LEI Pifeng, FANG Xi, DENG Xiangwen. Regulation mechanisms of forest vegetation restoration on water and soil erosion in mountainous and hilly area of Southern China[J]. Journal of Soil and Water Conservation, 2021, 35(5):1-9. doi: 10.13870/j.cnki.stbcxb.2021.05.001 [23] 陈保冬, 于萌, 郝志鹏, 谢伟, 张莘. 丛枝菌根真菌应用技术研究进展[J]. 应用生态学报, 2019, 30(3):1035-1046. doi: 10.13287/j.1001-9332.201903.037CHEN Baodong, YU Meng, HAO Zhipeng, XIE Wei, ZHANG Xin. Research progress in arbuscular mycorrhizal technology[J]. Chinese Journal of Applied Ecology, 2019, 30(3):1035-1046. doi: 10.13287/j.1001-9332.201903.037 [24] Zak R D, Pellitier P T, Argiroff W A, Castillo B, James T Y, Nave L E, Averill C, Beidler K V, Bhatnagar J, Blesh J, Classen A T, Craig M, Fernandez C W, Gundersen P, Johansen R, Koide R T, Lilleskov E A, Lindahl B D, Nadewlhoffer K J, Phillips R P, Tunlid A. Exploring the role of ectomycorrhizal fungi in soil carbon dynamics[J]. New Phytologist, 2019, 223(1):33-39. doi: 10.1111/nph.15679 [25] 郭屹立, 李冬兴, 王斌, 白坤栋, 向悟生, 李先琨. 北热带喀斯特季节性雨林土壤和6个常见树种凋落物的C、N、P化学计量学特征[J]. 生物多样性, 1085, 25(10):1085-1094.GUO Yili, LI Dongxing, WANG Bin, BAI Kundong, XIANG Wusheng, LI Xiankun. C, N and P stoichiometric characteristics of soil and litter fall for six common tree species in a northern tropical karst seasonal rainforest in Nonggang, Guangxi, Southern China[J]. Biodiversity Science, 25, 1085, 25(10):1085-1094. [26] 胡宁, 马志敏, 蓝家程, 伍宇春, 傅瓦利, 袁红, 娄露玲. 石漠化区植被恢复过程凋落叶分解特征及其对土壤有机碳/氮的影响:以重庆中梁山为例[J]. 中国岩溶, 2016, 35(5):539-549.HU Ning, MA Zhimin, LAN Jiacheng, WU Yuchun, FU Wali, YUAN Hong, LOU Luling. Leaf litter decomposition characters and impact on soil organic carbon/nitrogen in different vegetation restorations of karst rocky desertification: An example of the Zhongliang mountain in Chongqing[J]. Carsologica Sinica, 2016, 35(5):539-549. [27] Sluiter A, Hames B, Ruiz R, Scarlata C, Sluiter J, Termpleton D, Crocker D. Determination of structural carbohydrates and lignin in biomass[R]. Golden: National Renewable Energy Laboratory, 2008. [28] Hagerman A E. The Tannin Handbook[M]. Oxford: Miami University, 2011. [29] Cotrufo M, Soong J, Horton A J, Campbell E E, Haddix M L, Wall D H, PartonW J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss[J]. Nature Geoscience, 2015, 8:776-779. doi: 10.1038/ngeo2520 [30] 潘复静, 张伟, 王克林, 何寻阳, 梁士楚, 韦国富. 典型喀斯特峰丛洼地植被群落凋落物C∶N∶P生态化学计量特征[J]. 生态学报, 2011, 31(2):335-343.PAN Fujing, ZHANG Wei, WANG Kelin, HE Xunyang, LIANG Shichu, WEI Guofu. Litter C∶N∶P ecological stoichiometry character of plant communities in typical karst peak-cluster depression[J]. Acta Ecologica Sinica, 2011, 31(2):335-343. [31] 吴鹏, 崔迎春, 赵文君, 侯贻菊, 朱军, 丁访军, 杨文斌. 茂兰喀斯特区68种典型植物叶片化学计量特征[J]. 生态学报, 2020, 40(14):5063-5080.WU Peng, CUI Yingchun, ZHAO Wenjun, HOU Yiju, ZHU Jun, DING Fangjun, YANG Wenbin. Leaf stoichiometric characteristics of 68 typical plant species in Maolan National Nature Reserve, Guizhou, China[J]. Acta Ecologica Sinica, 2020, 40(14):5063-5080. [32] 曾昭霞, 王克林, 曾馥平, 宋同清, 刘孝利, 宋希娟. 桂西北喀斯特区原生林与次生林凋落叶降解和养分释放[J]. 生态学报, 2012, 32(9):2720-2728. doi: 10.5846/stxb201103240372ZENG Zhaoxia, WANG Kelin, ZENG Fuping, SONG Tongqing, LIU Xiaoli, SONG Xijuan. Litter decomposition and nutrients release in typical secondary and primary forests in karst region, northwest of Guangxi[J]. Acta Ecologica Sinica, 2012, 32(9):2720-2728. doi: 10.5846/stxb201103240372 [33] 田地, 严正兵, 方精云. 植物生态化学计量特征及其主要假说[J]. 植物生态学报, 2021, 45(7):682-713. doi: 10.17521/cjpe.2020.0331TIAN Di, YAN Zhengbing, FANG Jingyun. Review on characteristics and main hypotheses of plant ecological stoichiometry[J]. Chinese Journal of Plant Ecology, 2021, 45(7):682-713. doi: 10.17521/cjpe.2020.0331 [34] 唐仕姗, 杨万勤, 王海鹏, 熊莉, 聂富育, 徐振峰. 中国森林凋落叶N、P化学计量特征及控制因素[J]. 应用与环境生物学报, 2015, 21(2):316-322.TANG Shishan, YANG Wanqin, WANG Haipeng, XIONG Li, NIE Fuyu, XU Zhenfeng. Stoichiometric characteristics and controlling factors of N and P in forest leaf litter of China[J]. Chinese Journal of Applied and Environmental Biology, 2015, 21(2):316-322. [35] Chen H, Li D J, Xiao K C, Wang K L. Soil microbial processes and resource limitation in karst and non-karst forests[J]. Functional Ecology, 2018, 32(5):1400-1409. doi: 10.1111/1365-2435.13069 [36] Poorter L, Bongers F. Leaf traits are good predictors of plant performance across 53 rain forest species[J]. Ecology, 2006, 87(7):1733-1743. doi: 10.1890/0012-9658(2006)87[1733:LTAGPO]2.0.CO;2 [37] Chomel M, Guittonny-Larchevêque M, Fernandez C, Gallet C, DesRochers A, Pare D, Jackson B G, Baldy V, Aerts R. Plant secondary metabolites: A key driver of litter decomposition and soil nutrient cycling[J]. Journal of Ecology, 2016, 104(6):1527-1541. doi: 10.1111/1365-2745.12644 [38] Zhang H Y, Lu X T, Hartmann H, Keller A, Han X G, Trumbore S, Phillips R P. Foliar nutrient resorption differs between arbuscular mycorrhizal and ectomycorrhizal trees at local and global scales[J]. Global Ecology & Biogeography, 2018, 27(7):875-885. [39] Chen H, Li D J, Mao Q G, Xiao K C, Wang K L. Resource limitation of soil microbes in karst ecosystems[J]. Science of the Total Environment, 2019, 650(1):241-248. [40] Shi J C, Zhao B Y, Zheng S, Zhang X W, Wang X L, Dong W T, Xie Q J, Wang G, Xiao Y P, Chen F, Yu N, Wang E. A phosphate starvation response-centered network regulates mycorrhizal symbiosis[J]. Cell, 2021, 184(22):5527-5540. doi: 10.1016/j.cell.2021.09.030 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3292
- HTML浏览量: 1513
- PDF下载量: 104
- 被引次数: 0
